

A graptolite sampler, from an ancient Encyclopedia Britannica
Occasionally, life looks like it isn’t. In the eastern forests of North America and in a thin strip along the Pacific Northwest (but sadly not in Colorado), hidden in plain sight on tree trunks you can find the gracefully named elven script lichen, Graphis scripta. With a little imagination, the lichen looks like secret writing, not like an eruptive fungal-algal symbiont that specializes in cohabiting in tree bark.
In the 18th century, Linnaeus, the father of taxonomy, faced a similar biological dilemma. He found patterns in rock he suspected were chemical or geological formations that looked as if they had been alive, but actually had not. He called them “graptolites” (= rock writing), and gave the name to a variety of things may or may not have been alive. Over time, however, the term was co-opted by paleontologists for a group of strange fossils that very much had been alive. The only problem was, no one knew what the living parts of these things actually looked like — or how they might be related to anything alive today.
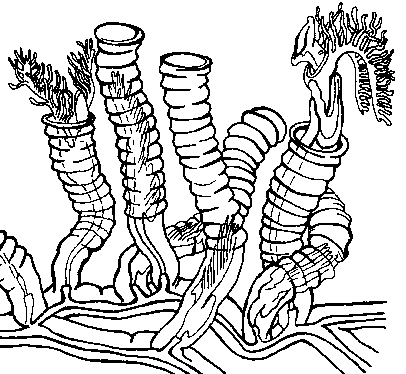
At left, you can see a variety of these creatures’s tube-like houses, called coenecia (se-nee-see-a), which are found abundantly through marine rocks from the Ordovician, Silurian, and part of the Devonian. Some were branched and tree like (dendritic, see13, 18, and 27, left) and probably bottom-dwellers, and others took on a variety of other bizarre forms that scientists interpret as the products of a planktonic form.
Many tended to have characteristic “hacksaw” shapes (see 4a, 7, 15, and 19, left) either in tuning forks, or coiled up in spirals like watch springs, as if something had poked out of the teeth lining these tubes. But no one knew what. The few cases where some actual ex-animal had fossilized were apparently more like ex-animal smudges than ex-animal fossils.

The pterobranch Rhabdopleura, in a lovely study in blue and gold. Note the tentaculate feeding/breathing prongs, aka lophophores.
Meanwhile, in another part of the science universe, scientists were describing and identifying members of a group called pterobranchs (= winged or feathered gills). Stretching little more than a centimeter long and living in proteinaceous* banded tubes cooperatively secreted by their shield-shaped probosci, they humbly go about their business stretching their ciliated tentaculate arms (which may remind you of bryozoans’lophophores, which they merely resemble convergently because that’s what they probably are (see comments)) into the water currents to catch prey and exchange gases. Inside their proboscis is a true lined body cavity, or coelom (seel-um). They sometimes live on their own, but usually grow in colonies connected by stems, or stolons**, in that colony of fused tubes called the coenecium. Some species have a pair of gill slits, just like fish (For a nice look at the general structure, see here, here, or, yes, the plush version here).
And this may very well reflect pterobranchs’ position in the shrub of life. Pterobranchs, it turns out, are hemichordates, in the same group as the acorn worms (enteropneusts) I described here last year. They are animals that evolved from animals just on the verge of becoming chordates, or nerve-corded animals like ourselves. They have a tripartite body plan of proboscis, collar (whence the tentaculate arms spring), and trunk. Like echinoderms (seastars, etc.) and chordates (us, etc.) they have a complete digestive tract whose mouth forms from the second indentation in the hollow ball of cells formed after a fertilized egg starts dividing (= deuterostome). Like echinoderms (but unlike chordates), they have no body segmentation and a special kind of larva called a dipleurula (chordates have no larvae). Signficantly, a hollow neural tube grows in some species early in development.
Most suggestively, pterobranchs and the fossil graptolites seemed suspicously similar, although little more than two dozen species of pterobranch live today, and for millions of years in the Paleozoic (from the Cambrian to the middle of the Devonian), graptolites were the dominant zooplankton in the world’s oceans. Tantalizingly, the microstructure of graptolite fossil tubes is very similar to the microstructure of pterobranchs, a detail discovered when electron microscopes first peered inside tubes of both animals and ex-animals in the 1970s. But no preserved fossil animal could confirm this.
Looks like a graptolite. Quacks like a graptolite. Could be a graptolite. Wouldn’t it be great if we had some soft tissues preserved in a graptolite fossil! Well, now we do.

Galeaplumosus, which was probably a two-armed model. The right arm is broken off, but two tentacles are still visible on it. "You don't look a day over 500 million years. You and Rhabdopleura could be sisters!" From Hou et al., Current Biology. Click for link.
In a March paper in Current Biology, scientists report the discovery of a tentaculate graptolite 525 million years old from the lower (early) Cambrian.
Finally, in all its glory, an animal poking out of a conical graptolite tube. And what an animal!For pterobranchs, they are, at shy of two inches (four centimeters), Yao-Ming-class. Which is fitting, because the fossil was found in China and dubbed Galeaplumosus abilus, from galea (helmet) and plumosus (feathered), and ab (away from) and nubilus (cloudy). Yunnan, where the fossil was found, means “south of the clouds”.
The fossil provides the clincher on graptolites’ true identities: a banded (probably secreted) cooperatively-made tube with contractile stalk and tentaculate feeding arms projecting from the opening is the M.O. of extant pterobranchs.
Looking carefully at the fossil, scientists were even able to discern possible cilia (silly-uh — little hairs that beat back and forth to draw in particles of food) on one tentacle, and a possible contractile stalk inside the shell. What scientists have, apparently, is the earliest, largest hemichordate animal (zooid) ever found, alive or dead, and it seems to show that their way of feeding and building a house have changed virtually not-at-all in 525 million years. Take that, sharks***.
The authors of the paper hypothesized that the rarity of specimens like this is probably a result of most graptolites’ planktonic lifestyle: on the long trip to the big sleep, most graptolites/pterobranchs probably decayed before they hit bottom, while the shell has proved decay-resistant in modern tests. That this animal was preserved, they suggest, means it was likely a bottom-dweller.
And that would fit with what we know about graptolite natural history. Scientists long suspected that the first graptolites, which tended to be tree-shaped (dendritic) and evolved in the Cambrian, were likely sessile bottom dwellers. Only later, in the next era, the Ordovician, did a floating planktonic form also emerge, the Earth’s first large zooplankton — and by far the dominant plankton of the early Paleozoic oceans. With their collaborative approach to constructing a floating colony, they were a bit like floating bee hives or wasp nests, if the wasps were all attached at the abodomen by stems, secreted their own cells (instead of building them from chewed up wood or mud), and never left the nest. Like the vast floating chains of colonial salps in today’s oceans (though much smaller), they must have been strange indeed.

The graptolite Pendeograptus fruticosus from the Lower Ordovician (477-474 mya) near Bendigo, Victoria, Australia. This style is referred to as the "tuning fork".
These planktonic co-ops evolved so abundantly and so quickly that they are commonly used as “index fossils” by the geology and petroleum geology sets to date rocks relative to each other with fine detail. In their heyday, thousands of species filled the oceans, common, widespread, quickly evolving and easily identifiable: a rock dater’s dream (errr . . . yeah. Ammonites have also been used this way.) In the Silurian, for instance, 40 different graptolite zones have been described, with an average duration of .7 million years — incredibly fine detail for geologic time, where dating anything to within a few tens of millions of years is usually considered spectacular.
Sadly, the planktonic graptolites went extinct in the middle Devonian, about 380-400 million years ago. Thus, the first (Galeaplumosus et al.) and last forms we find in the fossil record (from the mid-Cretaceous, near the end of the age of non-avian dinosaurs) are bottom-dwelling dendritic forms — as are the handful of species alive today, the humble survivors of a formerly world-dominating group****. But let us take the sunnier view. This post could have been titled “Graptolites Are Not Extinct!”.
_______________________________________________________
To learn more about graptolites and pterobranchs, see here, and a nice page here by the British and Irish Graptolite Group (BIG-G : ) ).
*made of a collagen-esque material, a family of animal proteins that help keep your muscles attached to your bones and your skin perky.
**A term for horizontal connections between organisms. Stolon, incidentally, is also the term botanists use for stems (*not* roots) that run along or jut below the ground from plant to plant (aka runners). If you’ve grown strawberries you have experienced this phenomenon.
***420-million year-old posers
**** much like brachiopods
![]()
Hou XG, Aldridge RJ, Siveter DJ, Siveter DJ, Williams M, Zalasiewicz J, & Ma XY (2011). An Early Cambrian Hemichordate Zooid. Current biology : CB PMID: 21439828
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{ 1 trackback }
{ 6 comments… read them below or add one }
Wait, pterobranchs are deuterostomes and they have a lophophore? Is this homologous to the protostome lophophore (and would the ancestral bilaterian have had one)?
Excellent question. I was suprised too, but this is what my biology text says, “Innovations in Feeding. Evolution in one lineage of deuterostomes resulted in several modifications of the lophophore and the coelomic cavity within it. These modifications provided new ways of capturing and handling food. All living representatives of this lineage (Hemichordates) are wormlike animals that live buried in marine sands or muds, under rocks, or attached to algae […] The ten living species of pterobranchs seem to have changed relatively little from the ancestors of their lineage […] In the other hemichordate lineage (the enteropneusts, or acorn worms — jf), the lophophore was apparently lost and the proboscis grew larger and became a digging organ . . . ” — Purves, Orians, and Heller. Life: The Science of Biology, Fourth edition.
It is possible they misinterpreted the structure and it is convergent (homoplasy), but the overall body plan *does* look a heck of a lot like a bryozoan (complete with u-shaped digestive track with anus opening near the mouth), even though these are deuterostomes and they are protostomes. Based on this and my book, I’d guess the common ancestor (the ancestral bilaterian) must have had it, and it was lost in all deuterostomes but the ones who have changed the least.
Anyone else want to weigh in? Thoughts welcome.
I dunno, a lot of sessile filter-feeders have a u-shaped gut. Clams and tunicates, to name one example each from protostomes and deuterostomes. It can imagine a long series of selective pressures favoring putting both the input and output ends of the digestive tract next to the cilia that are powering the flow of water and food and waste. And bilateral symmetry seems more useful for a worm than a sessile feeder.
you asked!
cool news about pterobranchs, though. It’s pretty neat to imagine a world ocean full of frondy zooplankton colonies!
Good points. I really don’t know, so that’s why I asked! I’ve qualifed it and I’ll change the text if someone can come up with evidence they’re definitely not lophophores. : )
They are definitely not actual lophophores. The similarity is striking (Halanych, 1993) but recent molecular phylogenies strongly suggest that the ancestral hemichordate was an enteropneust-like animal, not a pterobranch-like animal (for example, Cannon et al., 2009). I guess it just means that if you want to feed on little particles flowing past, there is pretty much one way to do it.
By the way, thanks for writing this article! I’m a graptolite worker and this exciting new find just sailed right past me with out me noticing.
-Dave Bapst, UChicago
Cannon, J. T., A. L. Rychel, H. Eccleston, K. M. Halanych, and B. J. Swalla. 2009. Molecular phylogeny of hemichordata, with updated status of deep-sea enteropneusts. Molecular Phylogenetics and Evolution 52(1):17-24.
Halanych, K. M. 1993. Suspension Feeding by the Lophophore-like Apparatus of the Pterobranch Hemichordate Rhabdopleura normanni. Biological Bulletin 185:417-427.
Excellent. This will get changed post-haste. Thanks for weighing in! Convergent evolution strikes again, it seems . . .